The effect of water on fat and mass gains by migratory warblers
Ido Tsurim and Nir Sapir
The following is a summary of a study aimed to examine the influence of water on the behavior and physiology of migratory warblers. A more detailed account of this study may be found in Sapir, N., Tsurim, I., Gal, B. & Abramsky, Z. 2004. The effect of water availability on fuel deposition of two staging Sylvia warblers. Journal of Avian Biology 35: 25-32.
Migratory flights require high energetic investments. The required energy is stored mostly as fat prior to the migratory flight, and is replenished along the migratory route during stop-over (staging) periods. Replenishing the fat reserves is especially important before crossing ecological barriers, such as large water bodies and deserts. However, not only the amount of fat stores counts, the speed in which it accumulates is important too. Fast accumulation of fat may shorten the stop-over duration, thus substantially shortening the overall migration time. This, in turn may highly improve the birdĺs survival and future reproduction.
Fat accumulation may be influenced by many factors. These may be related to the abundance and quality of the food at the stop-over site, the environmental conditions (such as predation and microclimatic conditions), and to the physiology of the bird itself. Drinking-water was suggested to improve food consumption and utilization in non-migratory and domestic birds.
 |
 |
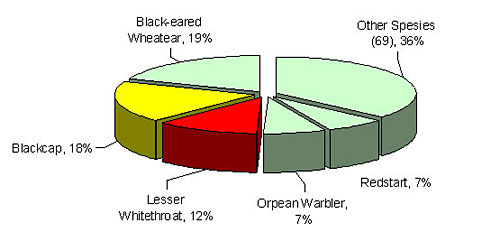
The goal of this study was to examine the effect of water availability on fat deposition of migratory passerines. Specifically, we manipulated water availability at a stop-over site and measured its effect on the density, fat deposition and body mass changes in blackcaps Sylvia atricapilla (picture 1) and lesser whitethroats S. curruca (picture 2) staging in a 3 ha plantation of Mount Atlas Gum-Tree Pistacia atlantica (pictures 3 and 5) located near Ein-Rimon, Northern Negev desert, Israel, during the autumns of 2000 and 2002. Many woodland passerine migrants concentrate in this site every autumn (Figure 1), just before they engage on a 1,800 km journey across the Sahara. The two species studied are abundant migrants in the area, and have long migratory periods, spanning between the beginning of August to the end of October. Both species were found to consume P. atlantica fruits (picture 4), with a 70% mean fruit volume in their feces. The birds digest the fruitĺs lipid-rich pulp (53 % of dry content) and excrete the seed. Water comprises only 34.5 % of the pulpĺs wet-content. Fruit abundance at the site ranged between 558,000 fruits during autumn 2000 and 1.5 million during autumn 2002. Yet, during the course of the study only about 30% of these fruits were ripe at any one time and thus appropriate for consumption by birds.
Experiment 1: In this experiment we examined the effect of water provisioning on the amount of fat accumulated by the studied species and on their activity level. It took place between 7 August and 19 October 2000. During this period we conducted 13 constant-effort bird-trapping sessions, using a 96 m mist-nets array. Each trapping session started at first light and lasted 4 hours. In two sessions (15 and 23 September) we provided the birds with drinking water. Water was provided approximately 20 hours before the start of the trapping effort. The remaining 11 trapping sessions, in which no water was provided, served as controls. Experimental trials were interspersed between control trials (e.g., 8, 19 and 25 September).
 |
Experiment 2: In this experiment we examined the effect of water provisioning on the rate in which fat was accumulated by individual birds. This was achieved by following body mass and fat changes of individual birds by re-trapping and measuring the same birds during a specific treatment session. The experiment was conducted during September 2002 in which we had a total of 17 constant-effort bird-trapping sessions, using a 141 m mist-nets array, again for the first 4 morning hours. Birds were trapped during three distinct periods (treatments), following each other. First, we trapped birds during 6 consecutive days (8 to 13 September), preceded by a single bird-trapping day (5 September). This period served as the first control trial and thus, water was not provided during this time. The second trapping period started immediately after the end of the first, and served as the water provision treatment. Water was provided throughout this period (14 to 18 September), starting 20 hours before the start of the first dayĺs bird trapping session. This treatment was immediately followed by a third trapping period (19 to 23 September) in which water was again unavailable to the birds. This last period served as a second control trial.
 |
Results Throughout both experiments we observed many birds, including warbler, drinking from the water provisioned at the site.
Experiment 1 (the effect of water on activity level, body mass and fat reserves): We trapped significantly more blackcaps during water provisioning days than during days with no water provisioning (Figure 2). This indicates that the overall activity level of blackcaps was higher when the stop-over site was supplemented with water. This may be the result of increased and prolonged activity of the blackcaps already on the site or from an increase in the blackcapsĺ density at the site. We have no means to decide which of the two mechanisms was more important.
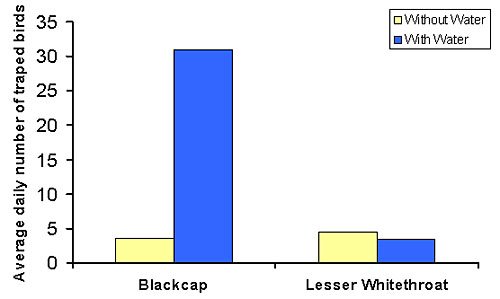
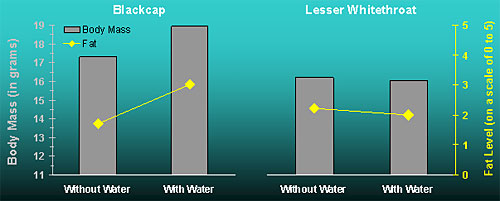
Additionally, the blackcaps captured during water provisioning sessions were heavier and had more fat reserves than blackcaps captured during days with no water supplementation (Figure 3). While this may indicate that blackcaps gained more mass and fat during water provisioning sessions, it is also possible that water attracted heavier and fatter blackcaps rather than induced higher body mass and fat gains. Again, have no means to decide which of the two mechanisms was more important.
Lesser whitethroats were not affected by the water supplementation (Figures 2 and 3).Throughout the study period we did not detect any relationship between changes in body mass and fat of the study species and the time along the migratory season. We therefore conclude that the body mass and fat changes observed during the study period were induced by the water provisioning.
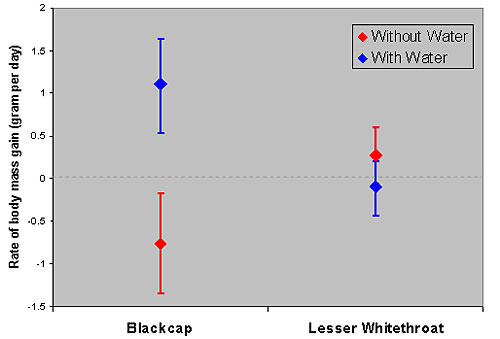
Experiment 2 (the effect of water on the rate of body mass and fat changes): We re-captured 26 warblers (15 Lesser Whitethroats and 11 Blackcaps), and calculated their rates of body mass and fat changes. Rates of change were calculated by subtracting the values at first capture from the values at last capture and then dividing the differences by the number of days passed between the two capturing events.
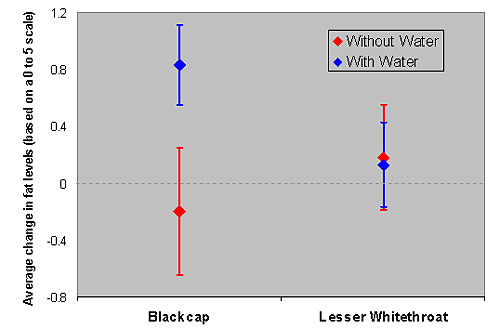
Blackcaps gained more body mass and fat during the water provisioning session in comparison to control sessions, in which water was not available at the study site. More importantly, the rates in which blackcaps accumulated body mass (Figure 4) and fat (figure 5) were higher during the water provisioning sessions. These findings support the hypothesis that water affects body mass and fat gains of blackcaps at the individual level and not only at the population level. I.e., the presence of water at the stop-over site does indeed improve body mass and fat gains by blackcaps.
As in Experiment-1 lesser whitethroats were not affected by the water supplementation (Figures 4 and 5)
Conclusions
Blackcaps gained more body mass and fat reserves and had done so faster when water was available, than during periods in which water was not available. The activity levels, and possibly the density, of blackcaps were also higher when water was available. However, water availability had no effect on body mass fat reserves and activity levels of lesser whitethroats. Yet, body mass and fat gains of lesser whitethroats, with or without water, were higher than those of blackcaps when water was not available. Species-specific differences in adaptations to arid conditions, reflected in the speciesĺ winter habitat preferences, may be responsible for the between-species dissimilarity in responding to water provision.
By facilitating mass and fat accumulation, water availability may influence habitat selection of long-distance migrants staging in arid and semi-arid areas along their migratory route. For staging birds that are sensitive to migration timing, gain of high water-induced energetic rewards during stopover may have an important influence on the birdsĺ migratory strategy with respect to energy and time savings, and may consequently determine the success of birds throughout their route and at their destination.
The results of this study indicate that water availability may have strong ecological and evolutionary consequences for birds migrating through arid environments, by its possible effect on bird behavior and physiology.
For additional details on the study site, additional readings on the subject, and related studies conducted at Ein Rimon, please consult:
Sapir, N. 2002. Stopover ecology of autumn migratory passerines in a man-made wood at a desert edge. - M. Sc. thesis, Ben-Gurion University.
Sapir, N., Tsurim, I., Gal, B. & Abramsky, Z. 2004. The effect of water availability on fuel deposition of two staging Sylvia warblers. Journal of Avian Biology 35: 25-32.
Shochat, E. 1999. Bird ringing in Be'er-sheva area and the northern Negev (in Hebrew with an English summary). - Torgos 28:34-40, 104.
Shochat, E., Abramsky, Z., Pinshow, B. and Whitehouse, M. E. A. 2002. Density-dependent habitat selection in migratory passerines during stopover: what causes the deviation from IFD? - Evol. Ecol. 16:469-488.
 |